Feuille
Cet article concerne l'utilisation du mot feuille en botanique. Pour l'utilisation du mot en papeterie, voir Feuille de papier. Pour les autres significations, voir Feuille (homonymie).
La feuille est, en morphologie végétale, l'organe spécialisé dans la photosynthèse chez les plantes vasculaires. Elle est insérée sur les tiges des plantes au niveau des nœuds. C'est aussi le siège de la respiration et de la transpiration. Les feuilles peuvent se spécialiser, notamment pour stocker des éléments nutritifs et de l'eau.


Pour accomplir son rôle, une feuille est généralement formée d'une lame plate et fine aérienne, le limbe, qui lui permet d'exposer à la lumière un maximum de surface. Mais il existe aussi des feuilles transformées, pour lesquelles le limbe est très réduit et ne joue plus de rôle photosynthétique ; elles sont transformées en vrilles, cataphylles, écailles sur les bourgeons, aériens (épines, aiguilles de conifères) ou souterrains (comme dans les bulbes, cormes), feuilles succulentes. C'est le parenchyme palissadique, un type particulier de tissu de la feuille, qui effectue la photosynthèse grâce à ses cellules contenant les chloroplastes, et donne à la feuille sa couleur verte. La feuille présente une grande variété de formes, de tailles, de teintes, de textures ou encore d'ornementations dans le règne végétal. Ces particularités de la feuille sont souvent caractéristiques d'une espèce, ou au moins d'un genre.
Les feuilles de certains légumes, tel le navet, sont appelés « fanes » ; d'autres feuilles comestibles sont des brèdes.
Le feuillage est constitué par l'ensemble des feuilles d'un arbre.
La feuillaison ou foliaison est un nom indénombrable qui désigne, pour les plantes annuelles, l'apparition des feuilles, phase saisonnière concomitante au débourrement.
Description
modifier




La figure 1 montre les différentes parties de la feuille : un limbe plan (a) parcouru de nervures (b), avec souvent un pétiole (c) qui rattache la feuille à la tige, parfois élargi en gaine (d). Celle-ci peut « embrasser » la tige comme chez les poacées. Le pétiole peut être absent, la feuille est alors dite sessile. Il peut parfois être ailé, ou muni à sa base de stipules plus ou moins développés. Au point d'insertion du pétiole et de la tige, se trouve un bourgeon axillaire.
À la différence du reste de l'appareil végétatif de la plante (racine et tige), la feuille présente en général une symétrie bilatérale et non axiale.
Forme
modifierLa feuille est dite simple si le limbe est entier, ou composée s'il est découpé en plusieurs petites feuilles : les folioles. Selon la disposition des folioles sur l'axe principal de la feuille ou rachis, il est dit que la feuille est :
- pennée, si les folioles sont disposées comme les barbes d'une plume ;
- imparipennée si leur nombre est impair (il y a une foliole terminale) ;
- paripennée dans le cas contraire ;
- trifoliolée s'il y a trois folioles (trèfle) ;
- palmée ou digitée si elles sont disposées comme les doigts de la main. Cependant les palmes, feuilles du palmier, qui sont en fait des feuilles entières (Chamaedorea metallica), qui ont parfois une forme pennée en forme de plume (dattier), ou encore costapalmée (Sabal palmetto) si la structure est intermédiaire entre la forme palmée et pennée (soit l'aspect général d'un éventail mais avec un axe central assez marqué);
- pédalée si chaque foliole est insérée sur la foliole voisine (hellébore) ;
La feuille peut être doublement composée ;
- bipennée si les folioles sont composées de foliolules, c'est le cas des palmiers Caryota ;
- tripennée si les folioles sont elles-mêmes composées.
La forme de la feuille (fig. 2) :
- cordiforme, en forme de cœur (a) ;
- ovale (b) ;
- lancéolée (c) ;
- palmée, en forme de paume (de main).
Les formes sont très diversifiées :
- falciforme (en forme de faux) ;
- auriculée (avec des oreillettes à la base du limbe : laiteron) ;
- sagittée (en forme de flèche) ;
- peltée (lorsque le pétiole s'insère au milieu du limbe : capucine, taro) ;
- décurrente le limbe se prolonge sur la tige par des ailes (ex. : bouillon blanc) ;
- perfoliée quand les oreillettes se soudent donnant l'impression que la tige traverse le limbe ;
- connée quand deux feuilles opposées sont soudées par le limbe (chèvrefeuille) ;
- etc.
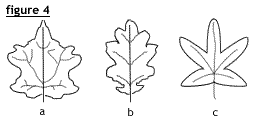
Division du limbe
modifierLe limbe peut être uni ou entier (comme fig. 1 et 2) ou plus ou moins profondément découpé (fig. 4). Dans ce dernier cas (fig.4), la feuille peut être :
- lobée (a), lorsque les lobes sont peu profonds et atteignent environ 1/8 de la largeur de la feuille, pennatilobée (b), lorsque les lobes sont disposés en peigne, palmatilobée lorsque les lobes ont une disposition palmée.
- pennatifide ou palmatifide, lorsque les divisions atteignent 1/4 de la largeur de la feuille et en tout cas moins de la moitié de celle-ci ;
- pennatipartite (c), ou palmatipartite lorsque les découpures dépassent le milieu de la feuille ;
- pennatiséquée (synonyme de composé pennée) ou palmatiséquée (composée palmée) si les divisions atteignent le rachis ;
- laciniée si le limbe est divisé en lanières étroites (plantes aquatiques).
-
 Différentes formes de feuilles
Différentes formes de feuilles -
 Différents aspects de bords de feuilles
Différents aspects de bords de feuilles -
 Différentes formes de feuilles découpées
Différentes formes de feuilles découpées



Bord du limbe
modifierLe bord du limbe (ou la marge) peut être uni ou entier (fig. 1 et 2), ondulé, sinué, scié (fig. 3a), serrulé (finement scié), denté (fig. 3b) ou crénelé (fig. 3c).
Disposition sur la tige
modifierEn outre, la distribution des feuilles sur la tige est aussi un caractère très variable, dont l'étude est la phyllotaxie. Elles peuvent être :
| Nom | Description | Illustration |
|---|---|---|
| alternes | isolées et disposées alternativement de part et d'autre de la tige (exemple : Saule à trois étamines) | 
|
| opposées | disposées par deux, au même niveau, l'un en face de l'autre sur une tige ou un rameau (exemple : le sorbier) | 
|
| distiques | disposées sur deux rangs opposés mais avec les limbes dans le même plan (exemples : l'if, orme champêtre) | 
|
| verticillées | réunies, par trois ou plus, en cercle autour de la tige, en étages successifs (exemple : Allamanda cathartica | 
|
| décussées | opposées et disposées à angle droit par rapport au nœud précédent ou suivant (exemple : lamier blanc) | 
|

Au sein d'une même espèce (chez les arbres notamment), les feuilles et les branches peuvent s'agencer de manière différente selon leur position dans l'arbre et leur exposition à la lumière. Ce phénomène participe de la morphologie générale des arbres, propre à chaque espèce.
Les feuilles excitées par le vent font scintiller et murmurer le feuillage. Les plantes sciaphiles de sous-bois et de forêts qui présentent généralement un indice foliaire élevé, donc un ombrage mutuel des feuilles très important, ont un feuillage qui forme un couvert discontinu, avec des trouées et des taches de soleil. Elles sont certainement adaptées à l'utilisation optimale des taches de soleil par le feuillage au vent, la disposition des feuilles et l'ouverture différentielle des stomates[1],[2]. Cette optimisation de la photosynthèse évite de saturer les feuilles de la partie supérieure de la canopée et de maintenir les feuilles de la partie inférieure dans l’ombre[3].
Polymorphisme foliaire
modifierLa forme des feuilles peut varier sur une même plante, c'est ce que l'on appelle l'hétérophyllie. C'est un phénomène assez courant dans le règne végétal.
C'est le cas chez le lierre grimpant où la forme des feuilles provenant de rameaux fertiles est différente de celle des feuilles appartenant aux rameaux stériles (polymorphisme vrai).
Le polymorphisme peut aussi résulter des conditions environnementales : chez la sagittaire à feuilles en flèche les feuilles immergées sont rubanées, les feuilles nageantes sont cordiformes, les feuilles aériennes sagittées.
Les cotylédons et les feuilles juvéniles qui leur succèdent immédiatement peuvent fréquemment être différentes de celles de l'âge adulte.
Plus généralement chez de nombreux végétaux, on retrouve des feuilles différentes (taille, forme) en fonction de la position de celles-ci sur l'individu. C'est par exemple le cas chez Morus alba.
Nombre
modifierLes plantes aphylles sont marquées par une forte réduction de la taille des feuilles, voire leur quasi-disparition ou leur disparition totale. À l'opposé, les grands pommiers ont de 50 à 100 000 feuilles, les bouleaux 200 000 en moyenne, les chênes à maturité 700 000 (mises côte à côte, ces 700 000 feuilles couvriraient une surface de 700 m2). Certains ormes d'Amérique ont à leur maturité jusqu'à cinq millions de feuilles. On estime que l'ensemble des feuilles des arbres du monde entier produisent par photosynthèse 65 000 à 80 000 millions de m3 de matière sèche par an, ce qui correspond aux deux tiers de la production mondiale des plantes terrestres[4].
Coloration
modifierLa coloration principale verte provient de la chlorophylle. Les feuilles juvéniles présentent parfois une teinte rougeâtre car le jeune tissu contient des anthocyanes de jeunesse, pigments bloquant les rayons ultraviolets, ce qui protège les petites feuilles des dommages de la photo-oxydation[5]. Le changement de couleur des feuilles montre l'existence de pigments complémentaires, les caroténoïdes, qui participent à la collecte des photons aux côtés des photosystèmes chlorophylliens. Les grandes variations dans la coloration des feuilles, pétioles et tiges, sans compter les fleurs et les fruits, ont une fonction aposématique : elles annulent le camouflage potentiel (i.e. l'homochromie) de certains insectes envers les oiseaux, par exemple[6].
L'observation à l'œil nu d'une feuille éclairée sous différents angles par le soleil laisse apparaître des taches brillantes blanches sur un fond vert. Cet effet optique, dû à la réflexion spéculaire de la lumière visible à la surface foliaire indépendante de la longueur d'onde, est plus ou moins marqué selon les espèces : certaines feuilles recouvertes d'une épaisse couche de cire apparaissent luisantes alors que d'autres tapissées de poils semblent plus ternes[7].
Composition chimique
modifierLa feuille est composée de pectine, de cellulose et de lignine. Ces composants sont de grandes molécules chimiques « emprisonnant » de nombreux éléments minéraux tels que calcium, potassium, sodium, magnésium, soufre, phosphore. Lors de la décomposition des feuilles en humus, ces éléments sont relâchés dans le sol et contribuent à son amélioration.
Reconnaissance des arbres par leurs feuilles
modifierLa feuille s'avère un critère relativement fiable pour identifier une plante ligneuse inconnue (arbre, arbrisseau ou grimpant ligneux).
Récemment en 2020, Régis Petit a proposé une méthode d'identification des arbres principalement par leurs feuilles. Les 8 critères principaux sont les suivants :
- plante (port) : arbre, arbrisseau ou grimpant ligneux ;
- plante (résine) : plante résineuse ou non ;
- plante (épines) : plante piquante ou non ;
- feuilles (type) : plate simple, plate composée, aiguille ou écaille ;
- feuilles (disposition sur tige) : isolées alternées, isolées opposées, groupées gainées ou groupées touffées ;
- feuille plate (squelette) : palmé ou penné ;
- feuille plate (bord) : denté ou lisse ;
- feuille plate (forme) : coeur, ronde, ovale, triangulaire/losangique, lancéolée, lobée ou à foliole composée.
Régis Petit a classé selon ces critères les 300 plantes ligneuses d'Europe les plus communes. Cette base de données est accessible gratuitement en ligne sous forme d'un fichier Excel multi-critères. L'identification d'une plante ligneuse inconnue en est ainsi facilitée (voir site référencé en Liens externes).
Fonctions
modifier



La fonction première des feuilles est la photosynthèse chlorophyllienne[10]. L'évolution des feuilles en organes de la photosynthèse s'est réalisée selon trois axes principaux[11] : l’exploitation de la lumière grâce au limbe plat qui présente une grande surface réceptrice[12], ce qui en fait un capteur solaire efficace[13] ; l’échange de gaz (CO2, O2 et évaporation d'H2O[14]) ; le transport de la sève brute et de la sève élaborée via un réseau très étendu de nervures. Selon le principe de l'allocation des ressources, les relations entre différents traits foliaires (forme, nervation, longueur du pétiole) reflètent l'existence de compromis évolutifs associés à des contraintes structurales et fonctionnelles des plantes en relation avec leur écologie[15].
Chez les plantes terrestres, les racines sont de loin la première source de nutriments (et parfois de contamination), mais les feuilles peuvent aussi absorber des nutriments autres que le CO2, voire le cas échéant des produits toxiques (ex : molécules de certains pesticides systémiques, particules métalliques fines et ultrafines issues de la pollution de l'air, dont métaux lourds et des métalloïdes) déposés à partir de l'air ou des pluies[16]. Ce transfert, qui semble se faire lentement au travers de la cuticule et/ou plus rapidement via les stomates est parfois un facteur déterminant du transfert des métaux de l'air vers la plante (Ceci a notamment été montré pour le plomb qui après s'être déposé sur les feuilles, est retrouvé dans les cellules situées sous la surface de la feuille)[17]. La morphologie des feuilles est l'un des facteurs qui favorise ou non le dépôt puis l'adsorption ou l'absorption et l'internalisation des particules atmosphériques. Il peut poser des problèmes de santé environnementale par exemple quand il concerne des légumes ou autres cultures alimentaires humaine ou à destination animale (fourrages...), surtout si le toxique est principalement stocké dans les parties comestibles (feuilles, fruits, graines, tubercules...). Bien comprendre ces processus est important pour le développement de l'agriculture urbaine (ou en zone polluée)[17]
Elles participent également à la défense des plantes contre les herbivores par synthèse de tanins, d'alcaloïdes ou de protéines PR, à la protection foliaire contre la photo-oxydation.
Les feuilles présentent toute une variété de spécialisation[18], dont par exemple :
- chez de nombreuses plantes aquatiques, les feuilles absorbent les nutriments dissous dans l'eau, parfois plus que les racines qui servent principalement à l'ancrage ;
- chez les plantes épineuses, les feuilles sont souvent transformées en épines issues de la modification des folioles, ou des stipules, ou simplement des poils. Comme les plantes xérophiles, il s'agit d'un mécanisme de défense contre la sécheresse, ou bien de défense contre le broutage des animaux herbivores. Certaines épines très fines, ainsi que des cils, permettent à la plante de collecter de la rosée (effet de pointe) ;
- chez les plantes carnivores, elles prennent des formes très spécialisées, en urne chez les Nepenthes, en piège chez les Dionées qui ont un limbe en deux parties munies d'aiguillons et capables de se replier l'une sur l'autre pour emprisonner des insectes ;
- chez les plantes grasses ou succulentes, les feuilles sont souvent transformées en organes de réserves ;
- chez les plantes grimpantes, les feuilles ou les folioles se transforment en vrilles leur permettant de s'accrocher à leur support. Parfois c'est le pétiole qui remplit cette fonction (Clématite) ;
- chez les plantes aquatiques, les feuilles peuvent se transformer en flotteurs (Jacinthe d'eau) ;
- chez les plantes xérophiles (adaptées à la sécheresse), les feuilles peuvent se réduire à des écailles (elles sont dites « squamiformes ») ou aiguilles (conifères). La plante diminue sa surface foliaire afin de limiter l'évapotranspiration. Ainsi le chêne vert peut avoir plusieurs formes de feuilles : en milieu favorable, où l'humidité de l'air n'est pas limitante, il aura des feuilles à limbe presque ovale, tandis qu'en milieu sec, les feuilles seront pour la plupart dentées.
- chez les épiphytes, certaines espèces se servent de leurs feuilles pour collecter et stocker de l'eau (Broméliacées par exemple)
Évolution
modifier


Il est possible qu'avant l'évolution des feuilles, les plantes aient développé la photosynthèse dans leurs tiges[note 2]. Les feuilles, actuellement, sont dans presque tous les cas une adaptation destinée à accroitre la quantité de lumière solaire utilisable pour la photosynthèse. La dichotomie microphylle/mégaphylle est aujourd'hui abandonnée pour expliquer l'histoire évolutive des feuilles qui sont certainement apparues une dizaine de fois indépendamment, et étaient probablement à l'origine des excroissances épineuses destinées à protéger les plantes primitives contre les herbivores[note 3], ce qui en ferait un exemple d'exaptation[19].
Les champignons endophytes sont une règle générale chez les feuilles (notion de mycophylle, feuille à endophyte fongique), cette stratégie symbiotique ayant certainement joué un rôle majeur dans le développement des feuilles au cours de l'évolution (rôle encore mal compris : amélioration de la résistance au stress hydrique, augmentation de la croissance et de la teneur en azote, réduction du broutage par les herbivores ?)[20].
Les feuilles sont actuellement les organes primaires de la photosynthèse chez les plantes. Elles sont classées en deux types : les feuilles simples, irriguées par une seule nervure et le plus souvent petites, et les feuilles composées, plus grandes et ayant une nervation complexe. Il a été suggéré que ces structures soient apparues indépendamment[21]. Les feuilles composées, selon la théorie du télome (de), auraient évolué à partir de branches présentant une architecture tridimensionnelle, à travers trois transformations : la planation, « aplatissant » cette architecture ; le tissage, formant des réseaux entre ces branches, et la fusion, regroupant ces réseaux pour former un limbe complet. Ces trois étapes auraient eu lieu à de multiples reprises dans l'évolution des feuilles modernes[22].
Les feuilles composées modernes sont probablement devenues majoritaires il y a 360 millions d'années, environ 40 millions d'années après la colonisation des terres au Dévonien inférieur par des plantes dépourvues de feuilles[23].
Cette évolution a été liée à la diminution de concentration du CO2 atmosphérique à la fin du Paléozoïque, elle-même associée à une augmentation de la densité des stomates à la surface des feuilles pour capter plus de CO2, ce qui assurait une meilleure évapotranspiration, et des échanges gazeux accrus[24]. La photosynthèse consommant moins de 5 % de l'eau apportée par la sève brute, les 95 % restant qui s'évapore favorisait un meilleur refroidissement des feuilles et leur permettait ainsi d'acquérir une plus grande surface[25],[26].
Les rhyniophytes de la flore de Rhynie n'étaient formés que de tiges minces et sans ornements. Aussi, les trimérophytes (en) du Dévonien moyen sont les premières plantes à présenter un aspect que l'on puisse qualifier de « feuillu ». Les plantes vasculaires de ce groupe se reconnaissent à des masses de sporanges situées à leurs extrémités, lesquelles peuvent bifurquer ou trifurquer[27]. Certains microphylles pourraient être ainsi des homologues de ces sporanges devenus stériles[28]. Certains organismes, tels que Psilophyton (en), portent des énations, de petites excroissances de la tige, épineuses ou poilues et dépourvues de vascularisation.
À peu près à la même époque, les zostérophyllophytes prenaient de l'importance. Ce groupe est reconnaissable à des sporanges en forme de rein, poussant sur de courtes branches latérales, à proximité de la tige principale, parfois ramifiées dans des formes en H caractéristiques[27]. La majorité des plantes de ce groupe portaient des épines sur leurs tiges, mais elles n'étaient pas vascularisées. Les premières traces d'énations vascularisées se trouvent dans le genre Asteroxylon. Les épines d'Asteroxylon présentent des traces de vaisseaux partant du protostèle central et allant irriguer chaque « feuille » individuelle. Un fossile connu sous le nom de Baragwanathia apparait un peu plus tôt, au Silurien supérieur[29] ; dans cet organisme, ces traces de vaisseaux continuent jusqu'au milieu de la feuille[30]. La théorie des énations soutient que les feuilles de type microphylle se sont développées comme des excroissances du protostèle se reliant à des énations déjà existantes, mais il est également possible que les feuilles simples aient évolué à partir de tiges se ramifiant et formant un réseau[27].
Asteroxylon[31] et Baragwanathia sont généralement considérés comme des lycopodes primitifs[27]. Les lycopodes existent toujours aujourd'hui (par exemple l’Isoète) ; ces lycopodes modernes portent des feuilles simples. Elles pouvaient être assez grandes – les Lepidodendrales avaient des feuilles simples de plus d'un mètre de long – mais elles sont presque toutes irriguées par un seul vaisseau, à l'exception de la ramification observée chez Selaginella.
Les feuilles composées ont sans doute des origines séparées ; elles sont de fait apparues indépendamment à quatre reprises, chez les fougères, les prèles, les progymnospermes et les plantes à graines[32]. Elles semblent provenir de branches ramifiées, qui se sont d'abord chevauchées, puis reliées entre elles, jusqu'à évoluer vers la structure typique d'un limbe foliaire[30]. Cette théorie des mégaphylles explique pourquoi la « lacune foliaire » laissée lorsque le pétiole se sépare de la branche ressemble à une ramification[30]. Dans chacun des quatre groupes où sont apparues des feuilles composées, ces feuilles ont subi une évolution rapide entre la fin du Dévonien et le début du Carbonifère, se diversifiant jusqu'à ce que les formes se stabilisent au milieu du Carbonifère[32].
La fin de cette diversification peut être attribuée à des contraintes de développement[32] mais une question reste ouverte : pourquoi les feuilles ont-elles mis si longtemps à apparaître ? Les plantes avaient conquis le sol depuis au moins 50 millions d'années avant que les feuilles composées apparaissent de manière significative. Cependant, de petites feuilles composées étaient déjà présentes dans le genre Eophyllophyton (en) au début du Dévonien – ce n'est donc pas en soi leur complexité qui explique le temps écoulé avant leur généralisation[33]. La meilleure explication donnée jusqu'à présent est que le taux de CO2 atmosphérique diminuait rapidement à cette époque – divisé par 10 durant le Dévonien[34]. Ceci est en relation avec une multiplication par 100 de la densité des stomates. Les stomates permettent à l'eau de s'évaporer des feuilles ; la faible densité des stomates au début du Dévonien avait pour conséquence que des feuilles trop grandes se seraient échauffées, et cette densité ne pouvait augmenter, car les tiges et les systèmes de racines primitifs ne pouvaient apporter d'eau assez vite pour soutenir un rythme d'évapotranspiration accru[35].
Les arbres à feuillage caduc sont une réponse à un autre inconvénient des feuilles. La croyance populaire selon laquelle c'est une adaptation au raccourcissement des jours en hiver est erronée : des arbres à feuillage persistant prospéraient au-delà des cercles polaires durant la période chaude de la fin du Paléocène[36]. La raison de la perte des feuilles en hiver la plus souvent retenue est la protection contre le vent et la neige, cette perte diminuant la surface totale de l'arbre. Ce mécanisme de perte saisonnière est apparu à plusieurs reprises, et se présente actuellement chez les ginkgoales, les pinophyta et les angiospermes[37]. Il est possible également que ce mécanisme soit une réponse à la pression des insectes : il est peut-être moins coûteux d'abandonner entièrement les feuilles en hiver que de continuer à investir des ressources dans leur réparation[38].
Anatomie
modifier
Pétiole
modifierLe pétiole (du latin petiolus : petit pied) est le pédoncule de la feuille, reliant son limbe à la tige. Ses faisceaux conducteurs présentent une symétrie bilatérale, ce qui indique la nature foliaire et non caulinaire du pétiole. Lorsqu'il est élargi jusqu'à remplacer la feuille dans sa fonction, il est question de phyllode. Une feuille sans pétiole, ou à pétiole très court, est dite sessile.
Quelques pétioles ont des fonctions spécifiques : chez la sensitive, il permet le mouvement des feuilles ; chez la châtaigne d'eau, il permet la flottaison de la plante.
Nervures
modifier.jpg)
Les nervures d'une feuille sont les prolongements du pétiole dans le limbe foliaire. La nervure principale et les nervures secondaires partent de la première. C'est au niveau des nervures, se détachant par leur relief bombé du reste du limbe, que se situent l'essentiel des tissus conducteurs de sève (xylème et phloème), organisés en faisceaux.
La disposition des nervures (ou nervation) varie selon les espèces ou les familles. Il existe trois grands types de nervation :
- les feuilles à nervation pennée (penninerves), dans laquelle une nervure principale, prolongeant le pétiole, partage le limbe en deux parties sensiblement identiques selon l'axe de symétrie et à partir de laquelle les nervures secondaires se détachent selon une disposition alterne ou opposée ;
- les feuilles à nervation palmée (palmatinerves) où plusieurs nervures, en nombre impair, se détachent du pétiole au point de contact avec le limbe (exemple : la feuille de vigne) ;
- et les feuilles à nervation parallèle (parallélinerves), dont les nervures sont parallèles, sans anastomoses entre elles. C'est le cas de la plupart des graminées (Poaceae), dont les feuilles sont généralement sans pétiole.
Limbe foliaire
modifierLe limbe est constitué de tissus végétaux.
L'épiderme protecteur recouvre les surfaces supérieure et inférieure du limbe (appelées aussi faces adaxiale et abaxiale) ; il est constitué le plus souvent par une couche unique de cellules ne comportant généralement pas de chloroplastes, parfois couverte par une couche protectrice externe, la cuticule. Certaines cellules de l'épiderme peuvent se transformer en poils. Sur l'épiderme inférieur se trouvent les stomates. Ce sont des sortes de pores, formé par deux cellules en forme de reins, qui laissent entre elles une ouverture variable, l'ostiole.
Le mésophylle, ou parenchyme foliaire, comporte deux couches : sous l'épiderme supérieur, un parenchyme palissadique, tissu formé de plusieurs rangées de cellules allongées perpendiculairement à la surface du limbe et serrées entre elles, sans lacunes. Entre celui-ci et l'épiderme inférieur un parenchyme lacuneux, à cellules plus grandes ménageant entre elles un réseau de lacunes, qui communique avec les stomates et assure les échanges gazeux avec l'extérieur.

Épiderme
modifierL'épiderme est la couche de cellules externes des feuilles. Cette couche est généralement transparente (ces cellules n'ont pas de chloroplastes) et couverte par une cuticule d'aspect cireux permettant de limiter les pertes en eau lors de trop fortes chaleurs. Chez les végétaux des climats secs cette cuticule est donc plus épaisse. La cuticule est parfois plus fine sur l'épiderme inférieur que sur l'épiderme supérieur.
L'épiderme inférieur est percé de pores appelés stomates. Ceux-ci permettent à l'oxygène et au dioxyde de carbone de rentrer et sortir des feuilles. La vapeur d'eau est aussi évacuée par les stomates au cours de la transpiration. Pour conserver de l'eau, les stomates peuvent se fermer pendant la nuit.
Des poils recouvrent l'épiderme de nombreuses espèces de plantes.
Mésophylle
modifierLa plus grande partie de l'intérieur d'une feuille, entre l'épiderme inférieur et supérieur, est composée d'un parenchyme appelé mésophylle. Ce tissu joue un rôle très important dans la photosynthèse.

Le mésophylle est composé de deux parties : vers la face supérieure, le parenchyme palissadique est constitué de cellules verticales, allongées et serrées, riches en chloroplastes : c'est dans ce parenchyme que se déroule l'essentiel de la photosynthèse. Vers la face inférieure se trouve le parenchyme lacuneux (ou spongieux), aux cellules plus arrondies et moins serrées. Les lacunes entre ces cellules contiennent les gaz échangés entre la feuille et l'atmosphère.
Chez les monocotyledones le mésophylle est homogène. Il est composé d'un parenchyme uniforme.
Phénologie
modifier
Les feuilles peuvent être persistantes (conservation du feuillage plusieurs années[39]), semi-persistantes (conservation de la majorité des feuilles bien que certaines soient remplacées à la belle saison) ou caduc (les feuilles de la plante ne durent que quelques mois puis tombent ; variante : marcescence) selon les espèces, les conditions climatiques et les saisons. Pour les végétaux non persistants, l'apparition des feuilles est appelé « feuillaison » ou « foliaison ».
La chute du feuillage caduc en automne s’accompagne d'un changement de couleurs variant du jaune au brun rouge et au rouge. En Amérique du Nord, la saison des couleurs génère une grande activité touristique surtout en Nouvelle-Angleterre et dans l'est du Canada.
Dépollution et phytoremédiation
modifierEn novembre 2010, des chercheurs ont avancé l'hypothèse selon laquelle les écosystèmes boisés feuillus seraient capables de mieux dépolluer l’air que ce qui était initialement pensé, pour les composés organiques volatils (COV) testés. Les expériences faites en laboratoire laissent penser que les feuilles absorbent même encore plus efficacement les COV et les détruisent (par conversion enzymatique) quand elles sont stressées par des blessures ou par certains polluants (de l'ozone et du méthyl vinyl cétone lors des expériences). Le cycle des COV oxygénés dans l'air devrait donc être revu et mieux incorporé dans les modèles globaux de chimie de l'atmosphère et de transport des polluants[40].
Quelques chiffres et records
modifierDes feuilles de Raphia regalis peuvent atteindre 25 m de long, 4 m de large et 100 kg, ce qui en fait les feuilles les plus longues du règne végétal[41]. Celles de Crassula connata (en) ont une taille qui peut être réduite à 1,3 mm[42].
Le nombre de feuilles des arbres est très variable, il dépend de leur âge, de leur diamètre et de leur hauteur. Ainsi un vieux hêtre pourpre de 80 ans qui a une hauteur de 25 m et un diamètre de 15 m possède 800 000 feuilles couvrant une surface de 1 600 m2, cet ensemble folaire consommant par jour 2,352 kg de dioxyde de carbone[note 4] (ce qui correspond à une consommation de 25 435 kJ ou encore un peu plus de 25,4 millions de joules, soit les besoins énergétiques journaliers de deux ou trois adultes) et produisant 1,6 kg de glucose et 1,712 kg d’oxygène par heure, ce qui couvre la consommation de dix hommes[43].
Notes et références
modifierNotes
modifier- Les feuilles et les pétales sont très minces (moins d’un millimètre). Contenant 90 % d'eau, elles devraient être translucides voire transparentes mais les lacunes aérifères facilitent la transmission du rayonnement vert. Cf Marc-André Selosse, Petites histoires naturelles. Chroniques du vivant, Actes Sud Nature, , p. 163
- Ce que font encore certaines plantes modernes, comme les plantes succulentes
- Pour plus de détails, on pourra consulter l'article défense des plantes contre les herbivores.
- Les racines consomment quant à elles 0,96 kg d’eau
Références
modifier- Sané de Parcevaux et Laurent Huber, Bioclimatologie. Concepts et applications, éditions Quae, , p. 282.
- (en) R. L. Chazdon, R. W. Pearcy, « The Importance of Sunflecks for Forest Understory Plants », BioScience, vol. 41, no 11, , p. 760–766 (DOI 10.2307/1311725).
- (en) John S. Roden, « Modeling the light interception and carbon gain of individual fluttering aspen (Populus tremuloides Michx) leaves », Trees, vol. 17, no 2, , p. 117–126 (DOI 10.1007/s00468-002-0213-3).
- (en) Peter A. Thomas, Trees. Their Natural History, Cambridge University Press, (lire en ligne), p. 13.
- Peter Wohlleben, La Vie secrète des arbres. Ce qu'ils ressentent. Comment ils communiquent, Les Arènes, (lire en ligne), p. 83.
- (en) Simcha Lev-Yadun, Gidi Ne'eman, « When may green plants be aposomatic ? », Biological Journal of the Linnean Society, vol. 81, no 3, , p. 413–416 (DOI 10.1111/j.1095-8312.2004.00307.x).
- [PDF] Stéphane Jacquemoud, Jean-Baptiste Féret & Susan L. Ustin, Compréhension et modélisation de la couleur des feuilles, École thématique interdisciplinaire du CNRS – Couleur, question d’échelle : l’espace, 2009, p. 186
- Cette propriété serait une stratégie cryptique (la feuille se confond avec l'ambiance lumineuses caractérisée par des taches de soleil) ou un effet visuel (elle apparaît moins grande aux yeux des prédateurs). Cf. (en) Peter D. Moore, Tropical Forests, Infobase Publishing, (lire en ligne), p. 119
- Peter D. Moore, op. cit., p.19
- Celle-ci est aussi assurée, dans une proportion moindre, par la tige, qui est généralement verte, et dont le parenchyme comporte des chloroplastes. Un bon exemple est le Genêt à balais, chez qui toute la tige est dite photosynthétique.
- William G. Hopkins, Physiologie végétale, De Boeck Supérieur, (lire en ligne), p. 143
- Cette surface est caractérisée pour les couverts végétaux par l’indice de surface foliaire.
- La surface réceptrice d’une feuille type est disposée à angle droit par rapport à la lumière incidente. La théorie de Monsi et Saeki prévoit que l’interception lumineuse dépende fortement de l’orientation des feuilles. cf (en) Masami Monsi, Toshiro Saeki, « On the factor light in plant communities and its importance for matter production », Annals of Botany, vol. 95, no 3, , p. 549–567 (DOI 10.1093/aob/mci052)
- La transpiration végétale consiste en une perte d'eau par évaporation à la surface des feuilles.
- (en) S. Lavorel et E. Garnier, « Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology », British Ecological Society, vol. 16, no 5, , p. 545-556 (DOI 10.1046/j.1365-2435.2002.00664.x)
- (en) Schreck, E., Foucault, Y., Sarret, G., Sobanska, S., Cécillon, L., Castrec-Rouelle, M.,... & Dumat, C. (2012) Metal and metalloid foliar uptake by various plant species exposed to atmospheric industrial fallout: mechanisms involved for lead. Science of the Total Environment, 427, 253-262.
- (en) TianTian Xiong, Annabelle Austruy, Antoine Pierart, Muhammad Shahid, Eva Schreck, Stéphane Mombo, Camille Dumat (2016) Kinetic study of phytotoxicity induced by foliar lead uptake for vegetables exposed to fine particles and implications for sustainable urban agriculture ; Journal of Environmental Sciences, Volume 46, pp. 16-27 (résumé)
- Aline Raynal-Roques, La botanique redécouverte, Belin, 1994, p. 204
- (en) Ashwani Kumar, Helena Fernández, Maria Angeles Revilla, Working with Ferns. Issues and Applications, Springer, , p. 80
- (en) George Carroll, « Fungal Endophytes in Stems and Leaves. From Latent Pathogen to Mutualistic Symbiont », Ecology, vol. 69, no 1, , p. 2-9 (DOI 10.2307/1943154)
- (en) Crane et Kenrick et Paul Kenrick, « Diverted development of reproductive organs : A source of morphological innovation in land plants », Plant System. And Evol., vol. 206, no 1, , p. 161–174 (DOI 10.1007/BF00987946)
- (en) Piazza P, et al., « Evolution of leaf developmental mechanisms », New Phytol., vol. 167, no 3, , p. 693–710 (PMID 16101907, DOI 10.1111/j.1469-8137.2005.01466.x)
- (en) Patricia G. Gensel, Henry Nathaniel Andrews, Plant life in the Devonian, Praeger, , p. 345
- (en) Osborne, C.P. Beerling, D.J. Lomax, B.H. Chaloner, W.G., « Biophysical constraints on the origin of leaves inferred from the fossil record », Proceedings of the National Academy of Sciences, no 101, , p. 10360-10362
- (en) Beerling D. et al., « Evolution of leaf-form in land plants linked to atmospheric CO2 decline in the Late Palaeozoic era », Nature, vol. 410, no 6826, , p. 352–354 (lire en ligne)
- (en) A perspective on the CO2 theory of early leaf evolution, analyse en ligne de cet article.
- (en) Stewart, W.N.; Rothwell, G.W., Paleobiology and the evolution of plants, Cambridge University Press, , p. 521pp
- (en) Paul Kenrick, « Palaeontology : Turning over a new leaf », Nature, vol. 410, , p. 309-310 (DOI 10.1038/35066649)
- (en) Rickards, R.B., « The age of the earliest club mosses : the Silurian Baragwanathia flora in Victoria, Australia », Geological Magazine, vol. 137, no 2, , p. 207–209 (DOI 10.1017/S0016756800003800, lire en ligne [abstract])
- (en) Kaplan, D.R., « The Science of Plant Morphology : Definition, History, and Role in Modern Biology », American Journal of Botany, vol. 88, no 10, , p. 1711–1741 (DOI 10.2307/3558347, JSTOR 3558347)
- (en) Taylor, T.N., « Perithecial ascomycetes from the 400 million year old Rhynie chert : an example of ancestral polymorphism », Mycologia, vol. 97, , p. 269–285 (PMID 16389979, DOI 10.3852/mycologia.97.1.269, lire en ligne [abstract])
- (en) Boyce, C.K., « Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants », Paleobiology, vol. 28, , p. 70–100 (DOI 10.1666/0094-8373(2002)028<0070:EODPAT>2.0.CO;2)
- (en) Hao, S., « Structure of the Earliest Leaves : Adaptations to High Concentrations of Atmospheric CO2 », International Journal of Plant Sciences, vol. 164, , p. 71–75 (DOI 10.1086/344557)
- (en) Berner, R.A., « Geocarb III : A Revised Model of Atmospheric CO2 over Phanerozoic Time », American Journal of Science, vol. 301, no 2, , p. 182 (DOI 10.2475/ajs.301.2.182, lire en ligne [abstract])
- (en) Beerling, D.J., « Evolution of leaf-form in land plants linked to atmospheric CO2 decline in the Late Palaeozoic era », Nature, vol. 410, no 6826, , p. 287–394 (PMID 11268207, DOI 10.1038/35066546)
- (en) Shellito, C.J., « Reconstructing a lost Eocene paradise : Part I. Simulating the change in global floral distribution at the initial Eocene thermal maximum », Global and Planetary Change, vol. 50, nos 1-2, , p. 1–17 (DOI 10.1016/j.gloplacha.2005.08.001, lire en ligne)
- (en) Aerts, R., « The advantages of being evergreen », Trends in Ecology & Evolution, vol. 10, no 10, , p. 402–407 (DOI 10.1016/S0169-5347(00)89156-9)
- (en) Labandeira, C.C., « Ninety-seven million years of angiosperm-insect association : paleobiological insights into the meaning of coevolution », Proceedings of the National Academy of Sciences of the United States of America, vol. 91, no 25, , p. 12278–12282 (PMID 11607501, PMCID 45420, DOI 10.1073/pnas.91.25.12278)
- Ces feuilles vivent en moyenne 3 ans.
- (en) T. Karl1, P. Harley, L. Emmons, B. Thornton, A. Guenther, C. Basu, A. Turnipseed et K. Jardine ; Report Efficient Atmospheric Cleansing of Oxidized Organic Trace Gases by Vegetation ; Online 21 October 2010 Science 5 November 2010: Vol. 330 no. 6005 p. 816-819 DOI: 10.1126/science.1192534 ; Résumé en anglais
- Francis Hallé, The longest leaf in palms ?, Principes 21: 18
- (en) Flora of North America, OUP USA, , p. 152.
- Daniel Model, « L'arbre est un symbole de puissance », Model Box, no 79, , p. 3 (lire en ligne)
Voir aussi
modifierBibliographie
modifier- Allen J. Coombes, Zsolt Debreczy, The book of leaves, The University of Chicago Press, The Ivy Press limited, 2010, accessible en français par la traduction et l'adaptation de Jérôme Goutier, sous le titre générique assez curieux Arbres. L'encyclopédie des 600 plus beaux arbres du monde, Flammarion, Paris, 2011, (ISBN 978-2-0812-5806-8)
Articles connexes
modifierLiens externes
modifier
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Voir un schéma détaillé de la structure d'une feuille.
- Projet collaboratif d'outil d'assistance à la reconnaissance d'espèces à partir de leurs feuilles (Pl@ntNet/Tela Botanica)
- (en) Manuel d'architecture des feuilles
- Aide à la reconnaissance des arbres principalement par leurs feuilles